北欧海夏季大风过程的阻热效应和散热效应
2012年在罗弗敦海盆布放了一套海气耦合浮标,其中2012年8月5日-27日共23天的数据非常完整,捕捉到了一次强南风过程和一次强北风过程。通过对大风期间海气热通量进行分析,给出了夏季北欧海暖流区海气相互作用的一般特征。
由于北大西洋暖流的存在,即使在夏季,北欧海暖流区的海温也高于气温,通常情况下海洋向大气释放感热和潜热。我们的观测表明,夏季南风将暖湿气流输送至北欧海。由于气温升高接近海温,海洋向大气的热传输较弱,感热释放被削弱。由于南风输送的水汽足够充足,抑制了海水蒸发,导致海洋潜热释放也被削弱。气温的升高导致大气有效回辐射增加,海洋损失的净长波辐射减少。因此,即使大风引起强烈的湍流混合,南风也引起感热、潜热和长波辐射大幅减小,海洋的总热通量不到60 W/m2。我们将其称为阻热效应。阻热效应期间海洋放热少而得到的短波辐射很强,导致海洋的热量消耗很少(图1)。
夏季北风将干燥的冷空气输送到北欧海,海气温差变大以及大风引起的强烈湍流作用可以促进海洋向大气传热,使海洋的感热释放迅速增加;干燥的空气有利于促进海水蒸发,强烈的湍流作用使海洋的潜热释放也迅速增加;空气中水汽含量少、气温低,导致大气回辐射减少,海洋损失的净长波辐射增加。北风期间海洋通过感热、潜热以及净长波辐射损失能量,我们将其称为散热效应。在本次浮标观测中,散热效应期间海面热通量合计达到240 W/m2,是阻热效应时的四倍以上。
由于夏季北风占比较少,北欧海暖流区主要以阻热效应为主。阻热效应期间海洋失热少而接收的热量多,夏季北欧海会储存大量能量,这些能量会随海流输送到北冰洋,起到加速北极变暖的作用。而到秋冬季节北风为主时则以散热效应为主,海洋向大气释放大量能量,是北欧海暖水的主要散热期。本项研究的研究结果适用于高纬度温暖的海洋,在北极圈内,北欧海是唯一有大量暖水的海域,对发生在这个海域的散热效应和阻热效应需要有更深入的认识。
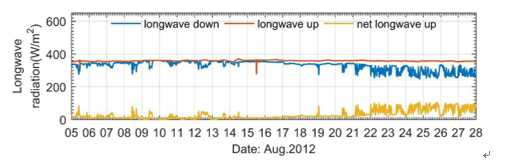


图1 观测期间两次风过程中长波辐射、感热和潜热通量的变化
三种国际主流的北极卫星遥感冰速产品评估和分析


图2 (a)-(e)分别代表北冰洋内区Ifremer-AMSR、OSISAF-AMSR、OSISAF-SSM、OSISAF-ASCAT、OSISAF-Merged相对IABP浮标冰速误差散点分布,红圈包含了均方根误差小于
1976-2018年北极固定冰时空变化特征分析
研究结果显示,北极固定冰主要出现在每年1-6月份,且分布于加拿大北极群岛的狭窄水道、东西伯利亚海沿岸、拉普捷夫海和喀拉海。20世纪80年代早期,北极固定冰年最大范围可达2.3 × 106 km2,至2018年减少了0.6 × 106 km2。1976-2018年间北极固定冰范围呈现出−1.1 ± 0.5 × 104 km2/yr (10.5%/decade)的显著减小趋势,相对变化率高于整个北极海冰的减小速率(5.2%/decade)。如图1b所示,2008-2018年间北极固定冰最大范围相比1976-2007年间减小了15%,从延伸序列来看,近十年来每年固定冰冬夏季范围均小于1976-2007年间的气候态平均值。
北极17个分区中仅白令海海域的固定冰范围及其持续时间呈现出同样的增加趋势。北极固定冰最大冰厚(>2.5 m)出现于加拿大北极群岛,冰厚增加速率也最高,达0.1 m/yr,该现象或许归因于此海域多年冰的输运与累积。而东北航道(图4)的固定冰平均厚度仅为1.57 m,且呈现出较大的空间变化性。其变化率(-1.2 cm/yr)远小于北极海冰的平均冰厚变化速度(-5.1 cm/yr)。平均而言,固定冰平均持续时间为44周,且以−0.06 ± 0.03 weeks/yr的速度持续减小。北极固定冰范围和冰厚减小速度与整体海冰相比均较小,这表明北极海冰剧烈衰减现象主要发生在浮冰区,而较大的相对范围损失指出北极固定冰区将更早更快的进入无冰状态。对北极沿岸地区固定冰的分析推动了对北极海冰变化的全面认识,同时得到了可用于北极航道研究的相关数据。

图3(a)1976-2018年北极固定冰范围年际变化;(b)2008-2018(新序列)年北极固定冰范围年际变化

图4 1976-2018年东北航道及西北航道固定冰范围变化
CMIP5和CMIP6 历史试验中BCC_CSM 北极海冰的模拟结果的评估
利用北京气候中心气候系统模式(BCC_CSM) 在最近两个耦合模式比较计划(CMIP5 和CMIP6) 的历史试验模拟结果,对北极海冰范围和冰厚的模拟性能进行了比较。结果表明:CMIP6改善了CMIP5模拟海冰范围季节变化过大的问题,总体上更接近观测结果;两个CMIP 试验阶段中BCC_CSM 模拟的海冰厚度都偏小,但CMIP6 试验对夏季海冰厚度过薄问题有所改进。通过对影响海冰生消过程的冰面和冰底热收支的分析认识到,8−9 月海洋热通量、向下短波辐射和反照率对模拟结果的误差影响较大,CMIP6 试验在这些方面有较大改善;而12 月至翌年2 月,CMIP5 模拟的北极海冰范围偏大主要是海洋热通量偏低所导致,CMIP6 模拟的海洋热通量较CMIP5 大,但北大西洋表层海流的改善才是巴芬湾附近海冰外缘线位置改善的主要原因。CMIP 试验模拟的夏季海冰厚度偏薄主要是因为6−8 月海洋热通量和冰面热收支都偏大,而CMIP6 试验模拟的夏季海冰厚度有所改善主要是由于海洋热通量和净短波辐射的改善(图5)。海冰模拟结果的改善与CMIP6 海冰模块和大气模块参数化的改进有直接和间接的关系,通过改变短波辐射、冰面反照率和海洋热通量,使BCC_CSM 模式对北极海冰的模拟性能也得到有效提高。

图5 6−8 月垂直方向总热收支、海洋热通量、冰面热收支、湍流通量、净辐射通量、净长波辐射、净短波辐射、反照率、向下短波辐射的分布
北极中央区经向水汽输送的主要通道
本研究采用两套再分析数据,分析了1979-2015年北极中央区的经向水汽输送,发现了水汽经向输送的主要通道。北极中央区有四条常年存在且较为稳定的水汽输送通道,分别位于拉普捷夫海、加拿大群岛和格陵兰岛两侧,它们在同一方向的输运频率皆超过了70%。此外,楚科奇海还存在一条仅在融冰-结冰期(3-9月)出现的季节性输送通道。通过以上通道的水汽占整个75oN纬圈上水汽输送的60%-80%。拉普捷夫海、楚科奇海和加拿大群岛这三条通道构成了较稳定的北极水汽通道。近年来北极太平洋扇区的经向水汽输送增加,融冰-结冰期尤为明显。波弗特高压的增强,使得通过北极太平洋扇区的水汽由净输出转为净输入(图6),相当于中央区每十年增加960km3液态水。而水汽经向输送的年际变化则主要出现在拉普捷夫海和东格陵兰岛的输送通道上。此外,融冰-结冰期北极中央区水汽输送的增加与5年冰面积的快速减少密切相关(相关系数达到-0.78),20世纪80年代以及最近十年两者几乎同步变化。


图6 a.1979-2015年北极大气经向水汽输送的气候态分布(NCEP-NCAR)及主要水汽通道,b.太平洋扇区三条通道的水汽输送强度变化(CSP—楚科奇海通道;LSP—拉普捷夫海通道;CAAP—加拿大北极群岛通道)
热带太平洋盐度变化:2015-2017盐度异常
本研究利用Argo海水盐度资料、海流同化数据和同期大气再分析数据,探讨了盐度趋势变化和相关动力过程。淡化海区的盐度异常集中在混合层内,从海区东部逐渐扩展到整个热带太平洋北部的大范围区域内;而咸化海区的盐度异常出现在温跃层附近,在异常事件中也具有显著的演变过程。2015年-2017年的盐度异常事件显著改变了上层盐度的长期变化(图7)。盐度平流和挟卷是造成这一盐度异常的主要动力因素,其中盐度平流的贡献最大,在淡化海区持续而显著,在咸化海区中后期的影响较为突出,体现为海洋环流的调整,另外次表层的挟卷过程也不可忽视。除此之外,影响盐度异常的因素还包括表层淡水通量和大尺度风场的驱动,次表层密度补偿引起的跨密度面混合也有着重要影响。盐度异常以及长期变化可以较好的反映全球气候变化的改变,也是衡量世界大洋气候变率的一个重要指标。
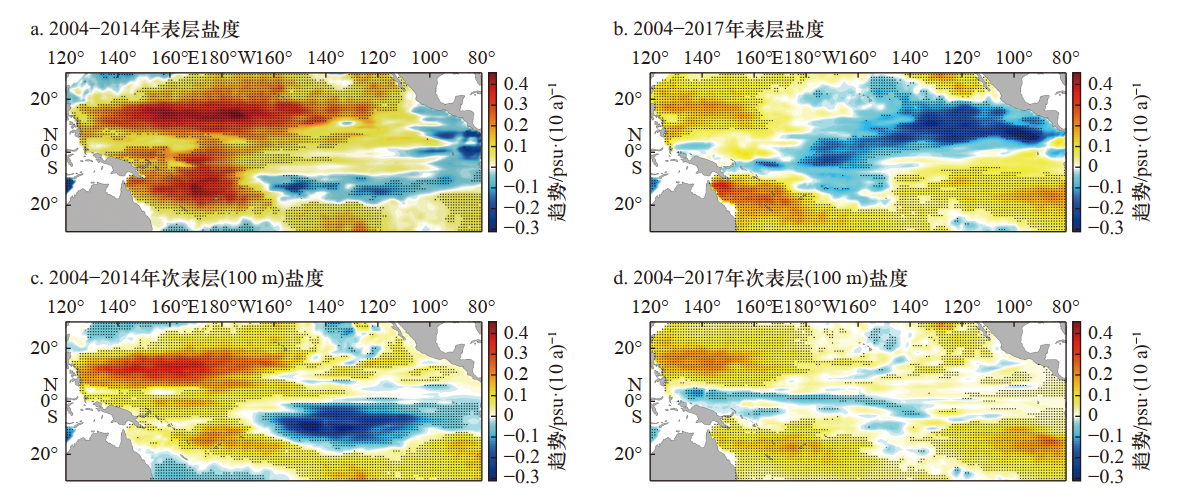

图7 2004-2014年和 2004-2017年热带太平洋表层和次表层(100 m)盐度线性变化趋势,以及两个时间段内上层盐度趋势之差的显著海域(e)
两极海冰对于全球表面温度非线性响应的贡献
本研究首次发现全球海表面温度(SST)对大小相同、符号相反的外强迫呈现出较强的非线性响应特征,表明了非线性是气候系统的固有特性。除此之外,本研究发现不论强迫来自于何处,冷强迫造成的降温总会强于热强迫造成的升温,且呈现出非常相似的空间分布,意味着我们目前的气候系统自身存在着一种趋冷的模态性响应(图9),因此,我们目前正在经历的全球升温极有可能是被非线性作用中和(减缓)之后的结果。非线性响应的量值在两极达到最大,尤其是在海冰边缘附近,且呈现出很强的季节变化特征。“温度归因法”分解的结果表明非线性的生成和反照率反馈和垂直递减率反馈息息相关。进一步地,通过在气候模式中抑制海冰的形成、去除季节变化,验证了海冰是导致非线性特征的根源,而季节变化作为“放大器”可以将其进一步放大。

图8 位于南纬55度的强迫激发出的(a)线性和(b)非线性响应。黑线为原始值,蓝线为无季节变化的情况,绿线为无海冰情况,红线为两者都无的情况。
北极高密集度冰区海冰的快速融化及其极端低值事件分析
利用美国冰雪中心(NSIDC)高分辨率海冰密集度等多种数据,定义了北极高密集度冰区(high concentration ice region: HCIR)海冰变化指数,在此基础上研究了1989-2017年HCIR海冰多尺度变化特征及其极端低值事件的可能形成原因。结果表明:北极HCIR海冰密集度具有显著的单峰型季节变化特征, 4月密集度最高,9月密集度最低,年较差达17.70%,兼有夏季融冰期短、冬季结冰期长且持续稳定的特点。HCIR海冰存在显著的年际年代际变化,在2007年发生了年代际转折以后,海冰变化指数的年际变化幅度和频次明显加强,且在2016、2012、2007、2011、2008和2010年依次出现海冰快速融化的密集度极端降低事件(图9a);2016年9月初HCIR海冰密集度达到历史最低值,接近50%。对HCIR海冰密集度极端低值事件的统计研究表明,29年间共出现874天(次)极端低值事件,约占总频次的8%;空间上海冰密集度的降低主要出现在沿HCIR边界线一带,存在巴伦支海-喀拉海北缘的斯瓦尔巴群岛-北地群岛和东西伯利亚-波弗特海两个中心区域(图9b),该空间分布与气旋式大气环流(图9c)引起的北冰洋Ekman漂流的辐散分布(图9d)相一致。这表明HCIR海冰密集度的极端降低,与极涡的动力作用有关,同时风场对海冰的动力辐散作用还会引起HCIR开阔水域的扩大,进一步加强海冰反照率的正反馈机制,使得热力和动力作用耦合起来共同影响HCIR海冰的加速融化。

图9 1989-2017年北极高密集度冰区海冰变化指数的逐日距平序列(a)(黑色:89-06年,红色:07-17年,图中蓝色虚线是89-17年的线性趋势线,黄色为89-06年线性趋势线,绿色为07-17年线性趋势线)以及基于北极高密集度冰区海冰密集度极端低值事件的夏季(7-9月)合成场。(b) 海冰密集度距平场; (c)表面2m气温距平场(填色等值线)、海平面气压场(黑色等值线)和850hPa 风场(矢量);(d)风生Ekman漂流(矢量)及其对应的散度场(填色等值线),其中图b-d中的绿色实线均代表北极高密集度冰区的范围边界
渤海海冰的年际变化及其预报研究
本文研究了渤海海冰的季节内和年际变化特征,分析了不同时间尺度冰情变化的影响因子,基于长短时记忆神经网络(LSTM)和可预报模态分析(PMA)法分别建立了延伸期和年尺度的冰情预报模型。首先通过合成分析和回归分析,研究了影响辽东湾海冰变化的局地和大尺度环流因子,并基于一种深度学习方法——长短时记忆神经网络(LSTM),建立了辽东湾海冰延伸期预报模型。结果表明(图10),LSTM模型能较好地预报出未来15d辽东湾海冰的总体发展趋势、浮冰外缘线离岸距离的振荡变化及峰值发生时间等关键特征,1-15d预报的平均绝对误差为4.1-5.7 海里,均方根误差为5.4-7.5海里。LSTM模型的预报时效可达到15d,较目前海冰数值预报(5-7d)的时效延长一倍,且运算速度极快,能够节省大量的计算资源和时间成本。该模型的建立为利用深度学习方法开展海洋和气象预报提供了一种新思路。
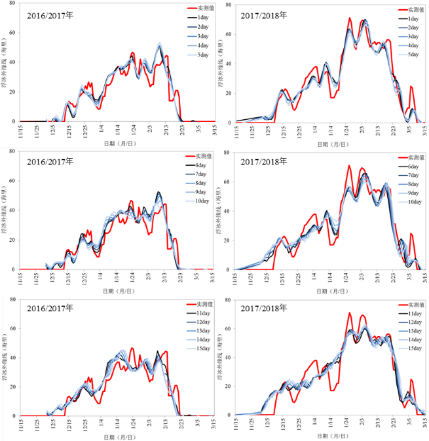
图10 LSTM的1~15 d预报结果与实测结果对比
北欧海低空大气的东西振荡模态对大气遥相关的“中继站”作用
北欧海上空的VP显示出除了表征北极涛动的全区一致型模态外,还表现出一个东西向辐合辐散反位相的东西振荡型模态(图11a,b)。该模态与一支连接北大西洋涛动(NAO)和欧亚遥相关(EU)的复合型大气遥相关波列有关(图11c),北欧海表层潜热和感热异常加热通过影响低空大气的异常辐合辐散,与北大西洋偶极子型海温异常加热共同作用,加强了NAO异常环流;同时高空急流波导作用(图11d,e)加强了从北欧海到东亚的EU波列,使得位于NAO和EU之间的北欧海成为连接NAO和EU大气遥相关波列的“中继站”,进而通过这种复合型大气遥相关波列将北大西洋与东亚大气环流联系起来,形成对东亚地区天气气候的远程影响。
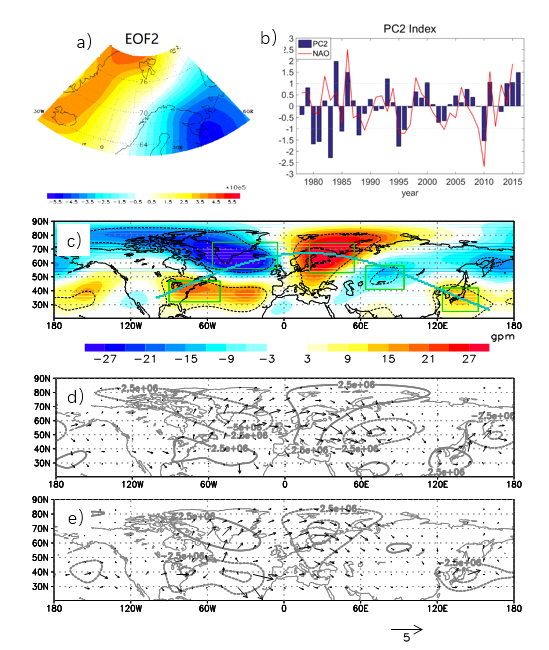
图11 北欧海秋冬季低层速度势EOF第二模态的空间场(a)与时间序列PC2(b)。(c)500hPa北半球位势高度对PC2的回归场,黑色虚线内为通过95%显著性检验的区域。(d)(e)PC2典型正(负)位相下的T-N波作用量(箭头)及对应的准地转流函数(等值线)
通过宏基因组学研究揭示了极地海洋微生物的结构与功能
本研究以极地地区28个站位采集到的60份海水样品(其中北极海域29份,南极海域21份)为研究对象,通过宏基因组学的研究手段与方法,系统表述了极地海洋微生物群落结构和代谢功能特点、极地与温带海域微生物群落结构的差异性。并在此基础上,通过对拼接获得的214个细菌基因组进行深入分析,对极地海洋微生物的基因组结构与功能及其适应极地环境的潜在机制进行了系统阐述。该研究成果对于完善全球海洋微生物组的认识具有十分重要的价值和作用。
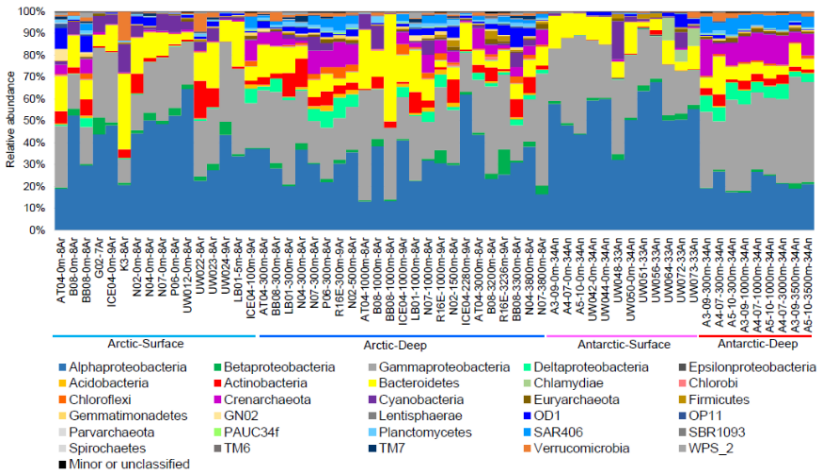
图12 极地海洋微生物群落结构组分相对丰度分布图(门级水平)
北极海洋细菌Polaribacter sp. SM1127胞外多糖能够促进大鼠皮肤伤口愈合和预防大鼠皮肤冻伤
研究了SM1127胞外多糖对大鼠皮肤的创面愈合促进和冻伤预防的作用。划痕实验表明,SM1127胞外多糖能促进人皮肤成纤维细胞的迁移。在Sprague-Dawley (SD)大鼠全层皮肤创伤实验中,通过宏观观察和组织学检查发现SM1127胞外多糖能够提高伤口愈合率,促进组织修复。在SD大鼠皮肤冻伤实验中,SM1127胞外多糖预处理大鼠皮肤可显著提高冻伤伤口愈合率,促进损伤皮肤的修复。这些结果表明SM1127胞外多糖能够促进皮肤伤口愈合和预防皮肤冻伤,在医药领域具有良好的应用潜力。

图13 SM1127胞外多糖对大鼠全层皮肤创伤伤口修复的影响
北极新型PL7亚家族褐藻胶裂解酶底物定位机制的结构与分子基础
该研究以实验室分离自北极褐藻Laminaria的一株冷单胞菌菌株分泌的一种新型褐藻酸裂解酶AlyC3为研究对象,基于对蛋白系统发生分析和酶学性质的研究,提出了建立多糖裂解酶7家族内一个以AlyC3为代表的新的亚家族,即亚家族6。该亚家族蛋白能够特异性降解甘露糖醛酸聚糖。此外,结构和生化分析表明,AlyC3是同源二聚体,代表了第一个二聚的内切型褐藻酸裂解酶结构。AlyC3的酶活力可被NaCl激活,并采用新型的盐激活机制,即盐度通过影响AlyC3的聚集状态来调节酶活性。这是AlyC3适应海水盐度的一种策略,与其他盐激活的褐藻酸裂解酶的激活机制不同。作者进一步解析了酶的失活突变体与甘露糖醛酸二糖的蛋白-底物复合物结构。通过对蛋白结构的分析和生化验证,揭示了AlyC3的催化过程。

图14 褐藻酸裂解酶AlyC3催化过程示意图
南极海绵共附生真菌Aspergillus insulicola HDN151418代谢产物
采用正相和反相硅胶柱色谱、Sephadex LH-20凝胶柱色谱、中压液相色谱(MPLC)以及高效液相色谱(HPLC)等现代分离纯化方法,从菌株SRDP-39中分得8个化合物,其中 3个为新颖的aspochracin类环三肽化合物(Sclerotiotides M-O,1-3)(图15),应用高分辨率质谱、核磁共振谱、紫外光谱、红外光谱、化学转化、Marfey法等方法手段和其理化性质确定了全部化合物的平面及立体结构。运用基于氢-氢,氢-碳偶合常数以及NOE相关信号的构象分析和Mosher法成功解决了已知化合物Sclerotiotide L(4)柔性侧链手性中心的立体构型。抗菌研究结果发现,该类化合物(Sclerotiotides M-N,1-2)对蜡状芽孢杆菌(Bacillus cereus)、副溶血性弧菌(Vibrio parahemolyticus)、草分枝杆菌(Mycobacterium phlei)以及耐甲氧西林金黄色葡萄球菌(methicillin-resistant coagulase-negative staphylococci)等菌株具有一定的抑制活性,为进一步研究开发新型抗菌药物提供了重要的药用先导结构类型。

图15 菌株SRDP-39中分离得到的化合物结构示意图
南极海绵来源真菌Penicillium sp. HDN151272次级代谢产物
对菌株Penicillium sp. HDN151272代谢产物进行研究,从该菌株中分离到3个新的苯醌类化合物ketidocillinones A-C (1−3),3个化合物均以烯醇-酮式相互转化的形式存在(图16)。生物活性测试表明,化合物Ketidocillinones B和C(化合物2,3)对Mycobactrium phlei, Pseudomonas aeruginosa, Methicillin Resistant Coagulase-Negative Staphylococci (MRCNS) and Bacillus cereus等多种不同细菌显示出抗菌活性,化合物2的MIC为1.56-12.50 µg/mL,化合物3的MIC为6.25 - 25.00 µg/mL。

图16 Penicillium sp.菌株分离得到的化合物结构示意图
南极新菌分离与鉴定研究
采用纯培养方法分离培养南极长城站和中山站12个站点土壤样品中的微生物,共获得482(代表菌株序列收录号为MW227555-MW237831)株细菌,其中细菌290株,放线菌192株。优势属为嗜冷杆菌属(Psychrobacter,12%)、假单胞属(Pseudomonas,10%)和节杆菌属(Arthrobacter,9%)。有七株菌的16S rDNA序列与已知序列相似性较低,为细菌新种(表1),全部完成新菌鉴定工作。
表1 分离菌株中筛选出的潜在新菌
Table 1. The potential novel species of isolated bacteria
